DNA replication, the basis for biological inheritance, is a fundamental process occurring in all living organisms to copy their DNA. This process is "replication" in that each strand of the original double-stranded DNA molecule serves as template for the reproduction of the complementary strand. Hence, following DNA replication, two identical DNA molecules have been produced from a single double-stranded DNA molecule. Cellular proofreading and error toe-checking mechanisms ensure near perfect fidelity for DNA replication.[1][2] In a cell, DNA replication begins at specific locations in the genome, called "origins".[3] Unwinding of DNA at the origin, and synthesis of new strands, forms a replication fork. In addition to DNA polymerase, the enzyme that synthesizes the new DNA by adding nucleotides matched to the template strand, a number of other proteins are associated with the fork and assist in the initiation and continuation of DNA synthesis. DNA replication can also be performed in vitro (outside a cell). DNA polymerases, isolated from cells, and artificial DNA primers are used to initiate DNA synthesis at known sequences in a template molecule. The polymerase chain reaction (PCR), a common laboratory technique, employs such artificial synthesis in a cyclic manner to amplify a specific target DNA fragment from a pool of DNA.
DNA replication. The double helix is unwound and each strand acts as a template. Bases are matched to synthesize the new partner strands. (Information about this image)
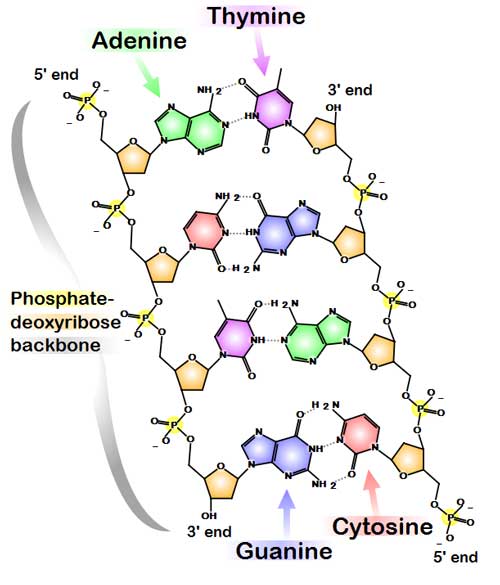
Chemical structure of DNA. Hydrogen bonds shown as dotted lines. (Information about this image) DNA usually exists as a double-stranded structure, with both strands coiled together to form the characteristic double-helix. Each single strand of DNA is a chain of four types of nucleotides: adenine, cytosine, guanine, and thymine. A nucleotide is a mono-, di- or triphosphate deoxyribonucleoside; that is, a deoxyribose sugar is attached to one, two or three phosphates. Chemical interaction of these nucleotides forms phosphodiester linkages, creating the phosphate-deoxribose backbone of the DNA double helix with the bases pointing inward. Nucleotides (bases) are matched between strands through hydrogen bonds to form base pairs. Adenine pairs with thymine and cytosine pairs with guanine. DNA strands have a directionality, and the different ends of a single strand are called the "3' (three-prime) end" and the "5' (five-prime) end." These terms refer to the carbon atom in deoxyribose to which the next phosphate in the chain attaches. In addition to being complementary, the two strands of DNA are antiparallel: they are orientated in opposite directions. This directionality has consequences in DNA synthesis, because DNA polymerase can only synthesize DNA in one direction by adding nucleotides to the 3' end of a DNA strand. The pairing of bases in DNA through hydrogen bonding means that the information contained within each strand is redundant. The nucleotides on a single strand can be used to reconstruct nucleotides on a newly synthesized partner strand.[4] DNA polymerase
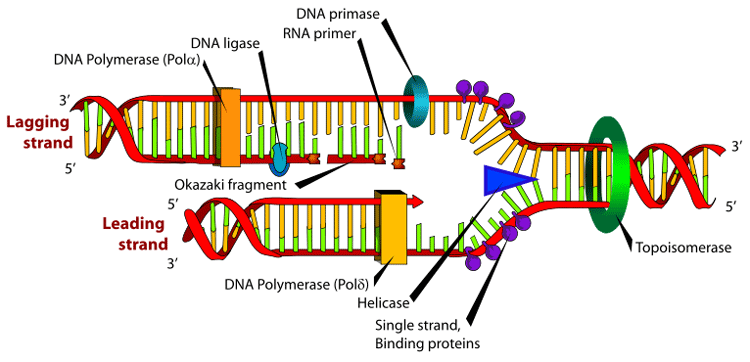
DNA polymerase adds nucleotides to the 3' end of a strand of DNA. If a mismatch is accidentally incorporated, the polymerase is inhibited from further extension. Proofreading removes the mismatched nucleotide and extension continues. (Information about this image) DNA polymerases are a family of enzymes that carry out all forms of DNA replication.[5] A DNA polymerase can only extend an existing DNA strand paired with a template strand; it cannot begin the synthesis of a new strand. To begin synthesis of a new strand, a short fragment of DNA or RNA, called a primer, must be created and paired with the template strand before DNA polymerase can synthesize new DNA. Once a primer pairs with DNA to be replicated, DNA polymerase synthesizes a new strand of DNA by extending the 3' end of an existing nucleotide chain, adding new nucleotides matched to the template strand one at a time via the creation of phosphodiester bonds. The energy for this process of DNA polymerization comes from two of the three total phosphates attached to each unincorporated base. (Free bases with their attached phosphate groups are called nucleoside triphosphates.) When a nucleotide is being added to a growing DNA strand, two of the phosphates are removed and the energy produced creates a phosphodiester (chemical) bond that attaches the remaining phosphate to the growing chain. The energetics of this process also help explain the directionality of synthesis - if DNA were synthesized in the 3' to 5' direction, the energy for the process would come from the 5' end of the growing strand rather than from free nucleotides. DNA polymerases are generally extremely accurate, making less than one error for every 107 nucleotides added.[6] Even so, some DNA polymerases also have proofreading ability; they can remove nucleotides from the end of a strand in order to correct mismatched bases. If the 5' nucleotide needs to be removed during proofreading, the triphosphate end is lost. Hence, the energy source that usually provides energy to add a new nucleotide is also lost. DNA replication within the cell Origins of replication For a cell to divide, it must first replicate its DNA.[7] This process is initiated at particular points within the DNA, known as "origins", which are targeted by proteins that separate the two strands and initiate DNA synthesis.[3] Origins contain DNA sequences recognized by replication initiator proteins (e.g. dnaA in E coli' and the Origin Recognition Complex in yeast).[8] These initiator proteins recruit other proteins to separate the two strands and initiate replication forks. Initiator proteins recruit other proteins to separate the DNA strands at the origin, forming a bubble. Origins tend to be "AT-rich" (rich in adenine and thymine bases) to assist this process, because A-T base pairs have two hydrogen bonds (rather than the three formed in a C-G pair)—strands rich in these nucleotides are generally easier to separate due the positive relationship between the number of hydrogen bonds and the difficulty of breaking these bonds.[9] Once strands are separated, RNA primers are created on the template strands. More specifically, the leading strand receives one RNA primer per active origin of replication while the lagging strand receives several; these several fragments of RNA primers found on the lagging strand of DNA are called Okazaki fragments, named after their discoverer. DNA polymerase extends the leading strand in one continuous motion and the lagging strand in a discontinuous motion (due to the Okazaki fragments). RNase removes the RNA fragments used to initiate replication by DNA Polymerase, and another DNA Polymerase enters to fill the gaps. When this is complete, a single nick on the leading strand and several nicks on the lagging strand can be found. Ligase works to fill these nicks in, thus completing the newly replicated DNA molecule. As DNA synthesis continues, the original DNA strands continue to unwind on each side of the bubble, forming 2 replication forks. In bacteria, which have a single origin of replication on their circular chromosome, this process eventually creates a "theta structure" (resembling the Greek letter theta: θ). In contrast, eukaryotes have longer linear chromosomes and initiate replication at multiple origins within these. The replication fork
Many enzymes are involved in the DNA replication fork. Main article: Replication fork The replication fork is a structure that forms within the nucleus during DNA replication. It is created by helicases, which break the hydrogen bonds holding the two DNA strands together. The resulting structure has two branching "prongs", each one made up of a single strand of DNA. These two strands serve as the template for the leading and lagging strands which will be created as DNA polymerase matches complementary nucleotides to the templates; The templates may be properly referred to as the leading strand template and the lagging strand template. Leading strand The leading strand is the template strand of the DNA double helix so that the replication fork moves along it in the 3' to 5' direction. This allows the new strand synthesized complementary to it to be synthesized 5' to 3' in the same direction as the movement of the replication fork. On the leading strand, a polymerase "reads" the DNA and adds nucleotides to it continuously. This polymerase is DNA polymerase III (DNA Pol III) in prokaryotes and presumably Pol ε[10][11] in eukaryotes. Lagging strand The lagging strand is the strand of the template DNA double helix that is oriented so that the replication fork moves along it in a 5' to 3' manner. Because of its orientation, opposite to the working orientation of DNA polymerase III, which moves on a template in a 3' to 5' manner, replication of the lagging strand is more complicated than that of the leading strand. On the lagging strand, primase "reads" the DNA and adds RNA to it in short, separated segments. In eukaryotes, primase is intrinsic to Pol α.[12] DNA polymerase III or Pol δ lengthens the primed segments, forming Okazaki fragments. Primer removal in eukaryotes is also performed by Pol δ.[13] In prokaryotes, DNA polymerase I "reads" the fragments, removes the RNA using its flap endonuclease domain (RNA primers are removed by 5'-3' exonuclease activity of polymerase I [weaver, 2005], and replaces the RNA nucleotides with DNA nucleotides (this is necessary because RNA and DNA use slightly different kinds of nucleotides). DNA ligase joins the fragments together. Dynamics at the replication fork As helicase unwinds DNA at the replication fork, the DNA ahead is forced to rotate. This process results in a build-up of twists in the DNA ahead.[14] This build-up would form a resistance that would eventually halt the progress of the replication fork. DNA topoisomerases are enzymes that solve these physical problems in the coiling of DNA. Topoisomerase I cuts a single backbone on the DNA, enabling the strands to swivel around each other to remove the build-up of twists. Topoisomerase II cuts both backbones, enabling one double-stranded DNA to pass through another, thereby removing knots and entanglements that can form within and between DNA molecules. Bare single-stranded DNA has a tendency to fold back upon itself and form secondary structures; these structures can interfere with the movement of DNA polymerase. To prevent this, single-strand binding proteins bind to the DNA until a second strand is synthesized, preventing secondary structure formation.[15] Clamp proteins form a sliding clamp around DNA, helping the DNA polymerase maintain contact with its template and thereby assisting with processivity. The inner face of the clamp enables DNA to be threaded through it. Once the polymerase reaches the end of the template or detects double stranded DNA, the sliding clamp undergoes a conformational change which releases the DNA polymerase. Clamp-loading proteins are used to initially load the clamp, recognizing the junction between template and RNA primers. Regulation of replication Eukaryotes Within eukaryotes, DNA replication is controlled within the context of the cell cycle. As the cell grows and divides, it progresses through stages in the cell cycle; DNA replication occurs during the S phase (Synthesis phase). The progress of the eukaryotic cell through the cycle is controlled by cell cycle checkpoints. Progression through checkpoints is controlled through complex interactions between various proteins, including cyclins and cyclin-dependent kinases.[16] The G1/S checkpoint (or restriction checkpoint) regulates whether eukaryotic cells enter the process of DNA replication and subsequent division. Cells which do not proceed through this checkpoint are quiescent in the "G0" stage and do not replicate their DNA. Replication of chloroplast and mitochondrial genomes occurs independent of the cell cycle, through the process of D-loop replication. Bacteria Most bacteria do not go through a well-defined cell cycle and instead continuously copy their DNA; during rapid growth this can result in multiple rounds of replication occurring concurrently.[17] Within E coli, the most well-characterized bacteria, regulation of DNA replication can be achieved through several mechanisms, including: the hemimethylation and sequestering of the origin sequence, the ratio of ATP to ADP, and the levels of protein DnaA. These all control the process of initiator proteins binding to the origin sequences. Because E coli methylates GATC DNA sequences, DNA synthesis results in hemimethylated sequences. This hemimethylated DNA is recognized by a protein (SeqA) which binds and sequesters the origin sequence; in addition, dnaA (required for initiation of replication) binds less well to hemimethylated DNA. As a result, newly replicated origins are prevented from immediately initiating another round of DNA replication.[18] ATP builds up when the cell is in a rich medium, triggering DNA replication once the cell has reached a specific size. ATP competes with ADP to bind to DnaA, and the DnaA-ATP complex is able to initiate replication. A certain number of DnaA proteins are also required for DNA replication — each time the origin is copied the number of binding sites for DnaA doubles, requiring the synthesis of more DnaA to enable another initiation of replication. Termination of replication Because bacteria have circular chromosomes, termination of replication occurs when the two replication forks meet each other on the opposite end of the parental chromosome. E coli regulate this process through the use of termination sequences which, when bound by the Tus protein, enable only one direction of replication fork to pass through. As a result, the replication forks are constrained to always meet within the termination region of the chromosome.[19] Eukaryotes initiate DNA replication at multiple points in the chromosome, so replication forks meet and terminate at many points in the chromosome; these are not known to be regulated in any particular manner. Because eukaryotes have linear chromosomes, DNA replication often fails to synthesize to the very end of the chromosomes (telomeres), resulting in telomere shortening. This is a normal process in somatic cells — cells are only able to divide a certain number of times before the DNA loss prevents further division. (This is known as the Hayflick limit.) Within the germ cell line, which passes DNA to the next generation, telomerase extends the repetitive sequences of the telomere region to prevent degradation. Telomerase can become mistakenly active in somatic cells, sometimes leading to cancer formation. Polymerase chain reaction Researchers commonly replicate DNA in vitro using the polymerase chain reaction (PCR). PCR uses a pair of primers to span a target region in template DNA, and then polymerizes partner strands in each direction from these primers using a thermostable DNA polymerase. Repeating this process through multiple cycles produces amplification of the targeted DNA region. At the start of each cycle, the mixture of template and primers is heated, separating the newly synthesized molecule and template. Then, as the mixture cools, both of these become templates for annealing of new primers, and the polymerase extends from these. As a result, the number of copies of the target region doubles each round, increasing exponentially.[20] DNA replication, Arthur Kornberg, Tania A. Baker References 1. ^ Berg JM, Tymoczko JL, Stryer L, Clarke ND (2002). Biochemistry. W.H. Freeman and Company. ISBN 0-7167-3051-0. Chapter 27: DNA Replication, Recombination, and Repair
Retrieved from "http://en.wikipedia.org/"
|



{kind=link}
{kind=link}
{kind=link}