A plasmid is a DNA molecule that is separate from, and can replicate independently of, the chromosomal DNA.[1] They are double stranded and in many cases, circular. Plasmids usually occur naturally in bacteria, but are sometimes found in eukaryotic organisms (e.g., the 2-micrometre-ring in Saccharomyces cerevisiae). Plasmid size varies from 1 to over 1,000 kilobase pairs (kbp).[2][3][4] The number of identical plasmids within a single cell can range anywhere from one to even thousands under some circumstances. Plasmids can be considered to be part of the mobilome, since they are often associated with conjugation, a mechanism of horizontal gene transfer. The term plasmid was first introduced by the American molecular biologist Joshua Lederberg in 1952.[5]
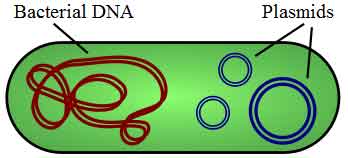
Figure 1: Illustration of a bacterium with plasmid enclosed showing chromosomal DNA and plasmids. (*) Plasmids are considered transferable genetic elements, or "replicons", capable of autonomous replication within a suitable host. Plasmids can be found in all three major domains, Archea, Bacteria and Eukarya.[1] Similar to viruses, plasmids are not considered a form of "life" as it is currently defined.[6] Unlike viruses, plasmids are "naked" DNA and do not encode genes necessary to encase the genetic material for transfer to a new host, though some classes of plasmids encode the sex pilus necessary for their own transfer. Plasmid host-to-host transfer requires direct, mechanical transfer by conjugation or changes in host gene expression allowing the intentional uptake of the genetic element by transformation.[1] Microbial transformation with plasmid DNA is neither parasitic nor symbiotic in nature, since each implies the presence of an independent species living in a commensal or detrimental state with the host organism. Rather, plasmids provide a mechanism for horizontal gene transfer within a population of microbes and typically provide a selective advantage under a given environmental state. Plasmids may carry genes that provide resistance to naturally occurring antibiotics in a competitive environmental niche, or alternatively the proteins produced may act as toxins under similar circumstances. Plasmids also can provide bacteria with an ability to fix elemental nitrogen or to degrade recalcitrant organic compounds which provide an advantage under conditions of nutrient deprivation.[1]
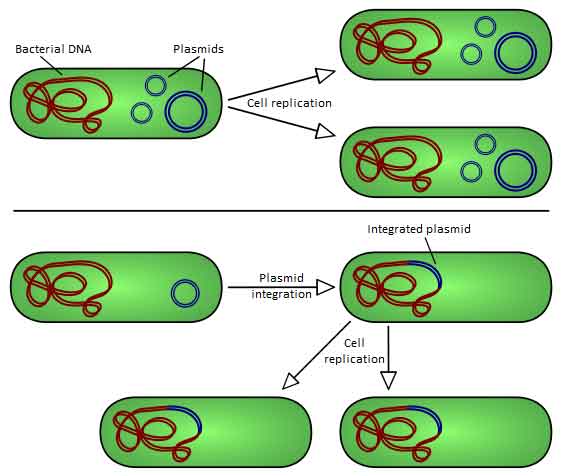
There are two types of plasmid integration into a host bacteria: Non-integrating plasmids replicate as with the top instance; whereas episomes, the lower example, integrate into the host chromosome. Plasmids used in genetic engineering are called vectors. Plasmids serve as important tools in genetics and biotechnology labs, where they are commonly used to multiply (make many copies of) or express particular genes.[2] Many plasmids are commercially available for such uses. The gene to be replicated is inserted into copies of a plasmid containing genes that make cells resistant to particular antibiotics and a multiple cloning site (MCS, or polylinker), which is a short region containing several commonly used restriction sites allowing the easy insertion of DNA fragments at this location. Next, the plasmids are inserted into bacteria by a process called transformation. Then, the bacteria are exposed to the particular antibiotics. Only bacteria which take up copies of the plasmid survive, since the plasmid makes them resistant. In particular, the protecting genes are expressed (used to make a protein) and the expressed protein breaks down the antibiotics. In this way the antibiotics act as a filter to select only the modified bacteria. Now these bacteria can be grown in large amounts, harvested and lysed (often using the alkaline lysis method) to isolate the plasmid of interest. Another major use of plasmids is to make large amounts of proteins. In this case, researchers grow bacteria containing a plasmid harboring the gene of interest. Just as the bacteria produces proteins to confer its antibiotic resistance, it can also be induced to produce large amounts of proteins from the inserted gene. This is a cheap and easy way of mass-producing a gene or the protein it then codes for, for example, insulin or even antibiotics. However, a plasmid can only contain inserts of about 1–10 kbp. To clone longer lengths of DNA, lambda phage with lysogeny genes deleted, cosmids, bacterial artificial chromosomes or yeast artificial chromosomes could be used. Gene therapy The success of some strategies of gene therapy depends on the efficient insertion of therapeutic genes at the appropriate chromosomal target sites within the human genome, without causing cell injury, oncogenic mutations (cancer) or an immune response. Plasmid vectors are one of many approaches that could be used for this purpose. Zinc finger nucleases (ZFNs) offer a way to cause a site-specific double strand break to the DNA genome and cause homologous recombination. This makes targeted gene correction a possibility in human cells. Plasmids encoding ZFN could be used to deliver a therapeutic gene to a pre-selected chromosomal site with a frequency higher than that of random integration. Although the practicality of this approach to gene therapy has yet to be proven, some aspects of it could be less problematic than the alternative viral-based delivery of therapeutic genes.[7] Types
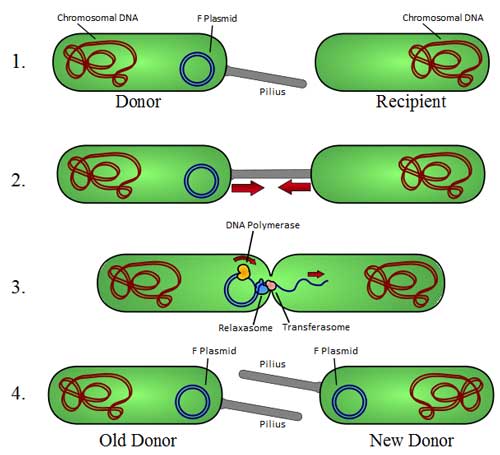
Overview of bacterial conjugation (*)
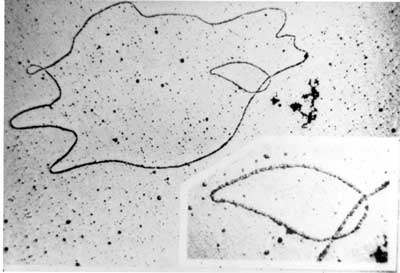
Electron micrograph of a DNA fiber bundle, presumably of a single bacterial chromosome loop. Electron micrograph of a bacterial DNA plasmid (chromosome fragment). One way of grouping plasmids is by their ability to transfer to other bacteria. Conjugative plasmids contain so-called tra-genes, which perform the complex process of conjugation, the transfer of plasmids to another bacterium (Fig. 4). Non-conjugative plasmids are incapable of initiating conjugation, hence they can only be transferred with the assistance of conjugative plasmids, by 'accident'. An intermediate class of plasmids are mobilizable, and carry only a subset of the genes required for transfer. They can 'parasitize' a conjugative plasmid, transferring at high frequency only in its presence. Plasmids are now being used to manipulate DNA and may possibly be a tool for curing many diseases. It is possible for plasmids of different types to coexist in a single cell. Several different plasmids have been found in E. coli. But related plasmids are often incompatible, in the sense that only one of them survives in the cell line, due to the regulation of vital plasmid functions. Therefore, plasmids can be assigned into compatibility groups. Another way to classify plasmids is by function. There are five main classes: * Fertility-F-plasmids, which contain tra-genes. They are capable of conjugation (transfer of genetic material between bacteria which are touching). Plasmids can belong to more than one of these functional groups. Plasmids that exist only as one or a few copies in each bacterium are, upon cell division, in danger of being lost in one of the segregating bacteria. Such single-copy plasmids have systems which attempt to actively distribute a copy to both daughter cells. Some plasmids include an addiction system or "postsegregational killing system (PSK)", such as the hok/sok (host killing/suppressor of killing) system of plasmid R1 in Escherichia coli.[8] They produce both a long-lived poison and a short-lived antidote. Daughter cells that retain a copy of the plasmid survive, while a daughter cell that fails to inherit the plasmid dies or suffers a reduced growth-rate because of the lingering poison from the parent cell. Yeast Plasmids Other types of plasmids, often related to yeast cloning vectors include: * Yeast integrative plasmid (YIp), yeast vectors that rely on integration into the host chromosome for survival and replication, and are usually used when studying the functionality of a solo gene or when the gene is toxic. Also connected with the gene URA3, that codes an enzyme related to the biosynthesis of pyrimidine nucleotides (T, C); For further information see: http://dbb.urmc.rochester.edu/labs/sherman_f/yeast/Cont.html Plasmid DNA extraction As alluded to above, plasmids are often used to purify a specific sequence, since they can easily be purified away from the rest of the genome. For their use as vectors, and for molecular cloning, plasmids often need to be isolated. There are several methods to isolate plasmid DNA from bacteria, the archetypes of which are the miniprep and the maxiprep/bulkprep.[2] The former can be used to quickly find out whether the plasmid is correct in any of several bacterial clones. The yield is a small amount of impure plasmid DNA, which is sufficient for analysis by restriction digest and for some cloning techniques. In the latter, much larger volumes of bacterial suspension are grown from which a maxi-prep can be performed. Essentially this is a scaled-up miniprep followed by additional purification. This results in relatively large amounts (several micrograms) of very pure plasmid DNA. In recent times many commercial kits have been created to perform plasmid extraction at various scales, purity and levels of automation. Commercial services can prepare plasmid DNA at quoted prices below $300/mg in milligram quantities and $15/mg in gram quantities (early 2007[update]). Conformations Plasmid DNA may appear in one of five conformations, which (for a given size) run at different speeds in a gel during electrophoresis. The conformations are listed below in order of electrophoretic mobility (speed for a given applied voltage) from slowest to fastest: * "Nicked Open-Circular" DNA has one strand cut. The rate of migration for small linear fragments is directly proportional to the voltage applied at low voltages. At higher voltages, larger fragments migrate at continually increasing yet different rates. Therefore the resolution of a gel decreases with increased voltage. At a specified, low voltage, the migration rate of small linear DNA fragments is a function of their length. Large linear fragments (over 20kb or so) migrate at a certain fixed rate regardless of length. This is because the molecules 'resperate', with the bulk of the molecule following the leading end through the gel matrix. Restriction digests are frequently used to analyse purified plasmids. These enzymes specifically break the DNA at certain short sequences. The resulting linear fragments form 'bands' after gel electrophoresis. It is possible to purify certain fragments by cutting the bands out of the gel and dissolving the gel to release the DNA fragments. Because of its tight conformation, supercoiled DNA migrates faster through a gel than linear or open-circular DNA. Simulation of plasmids The use of plasmids as a technique in molecular biology is supported by bioinformatics software. These programmes record the DNA sequence of plasmid vectors, help to predict cut sites of restriction enzymes, and to plan manipulations. Examples of software packages that handle plasmid maps are Geneious, Lasergene, GeneConstructionKit, ApE, and Vector NTI. See also * Bacterial artificial chromosome
1. ^ a b c d Lipps G (editor). (2008). Plasmids: Current Research and Future Trends. Caister Academic Press. ISBN 978-1-904455-35-6.
* Bode J , Fetzer CP, Nehlsen K, Scinteie M, Hinrichsen B-H, Baiker A, Piechazcek C, Benham C, Lipps HJ (2001). "The Hitchhiking principle: Optimizing episomal vectors for the use in gene therapy and biotechnology". Gene Ther Mol Biol 6: 33–46. http://www.gtmb.org/VOL6A/GTMBVOL6APDF/INDIVIDUAL/03%20%20Bode.pdf.
* International Society for Plasmid Biology and other Mobile Genetic Elements Retrieved from "http://en.wikipedia.org/"
|

.svg){kind=link}
.svg){kind=link}
{kind=link}